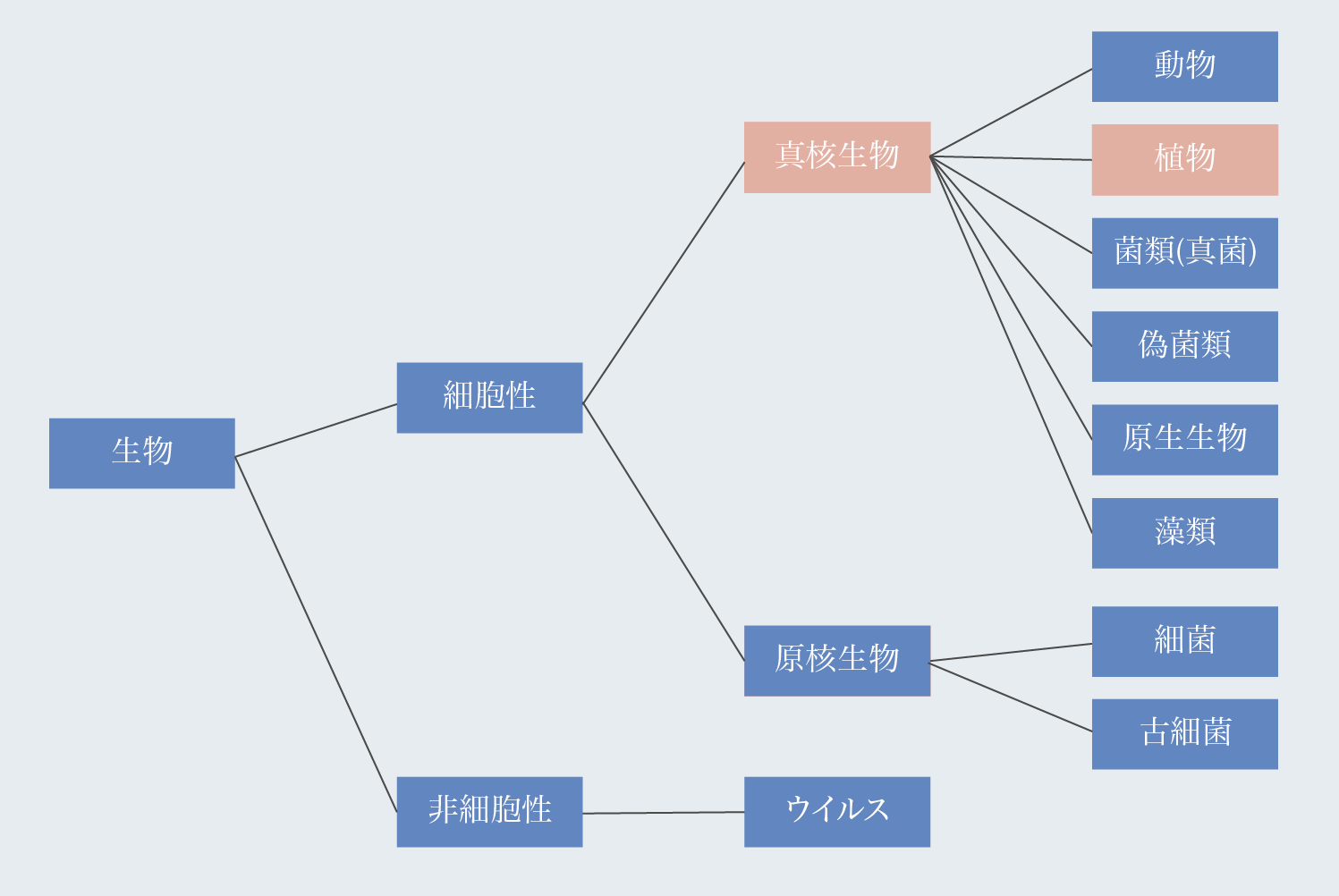
界(かい、英: kingdom、羅: regnum)は、生物学におけるドメインに次いで2番目に高い分類階級である。界は門と呼ばれるより小さなグループに分けられる。
米国やカナダの教科書では六界(動物界、植物界、菌界、原生生物界、古細菌界、細菌/真正細菌)が使われるのに対し、日本、イギリス、パキスタン、バングラデシュ、インド、ギリシャ、ブラジルなどの教科書では五界(動物界、植物界、菌界、原生生物界、モネラ界)が使われている。
現代分岐学に基づいた最近の分類の中には、従来の一部の界が単系統ではない、つまり共通の祖先のすべての子孫から構成されているわけではないことを指摘し、界という用語を明確に放棄しているものもある。また、植物相(flora)、動物相(fauna)、そして21世紀には真菌類(funga)という用語も、特定の地域や時代に存在する生命を表すのに使われる。
定義と関連用語
1735年、カール・リンネが、生物学に階級に基づく命名法を導入したとき、最も高い階級に「界」という名称を与え、その後に「綱」「目」「属」「種」という4つの主要な階級が続いた。その後、さらに2つの主要な階級が導入され、界(kingdom)、門(phylum, or division)、綱(class)、目(order)、科(family)、属(genus)、種(species)という順序になった。1990年、界の上位にドメイン(domain)という階級が導入された。
接頭辞を追加することができ、亜界(subkingdom、subregnum)と下界(infrakingdom、infraregnum)は、界(kingdom)の直下にある2つの階級である。上界(superkingdom)はドメインや帝国(empire)に相当するもの、あるいは帝国とドメインやサブドメインの中間に位置する独立した階級と考えることができる。分類体系によっては、亜界と下界の間に、さらに枝(branch、羅: ramus)の階級が挿入されることもある、たとえば、キャバリエ=スミスの分類における前口動物(Protostomia)や後口動物(Deuterostomia)があげられる。
歴史
生命の二界
生物を動物と植物に分類することは古くから行われてきた。アリストテレス(紀元前384-322年)は『History of Animals(動物誌)』で動物の種を分類し、弟子のテオプラストス(紀元前371 - 287年頃)は植物について『Historia Plantarum(植物誌)』を記述した。
1735年、カール・リンネ(1707-1778)は、現在では命名規約によって規定されている近代的な生物学的命名法の基礎を築いた。彼は、生物を2つの界「動物界」(Regnum Animale)と「植物界」(Regnum Vegetabile)に分けた。リンネはまた、分類体系に鉱物も含め、それらを第3の界「鉱物界」(Regnum Lapideum)に位置づけた。
生命の三界
1674年、しばしば「顕微鏡の父」と呼ばれるアントニ・ファン・レーウェンフックは、初めて顕微鏡で単細胞生物を観察した結果をロンドン王立協会に送った。それまでは、このような微生物の存在はまったく知られていなかった。けれどもリンネは彼の独創的な分類法に微生物を含めなかった。
当初、微生物は動物界と植物界に分類されていた。しかし、19世紀半ばには「植物界と動物界という既存の二分法は急速にその境界が曖昧になり、時代遅れになった」ことは、誰の目にも明らかになった。
1860年、ジョン・ホッグは、「すべての下等生物、あるいは原始的な有機生物」からなる第3の生命界である原生生物(Protoctista)を提案した。彼は、第4の界である鉱物界(Regnum Lapideum)を残した。1866年、エルンスト・ヘッケルもまた、動物でも植物でもない「中立的な生物」あるいは「原始的な形態の界」として、第3の生命界である原生生物界(Protista)を提唱した。しかし、ヘッケルはこの枠組みに鉱物界を含めなかった。ヘッケルは、生物が単細胞(原生生物界)か多細胞(動物界と植物界)かに基づく分類に落ち着くまで、この界の内容を何度も修正した。
四界
顕微鏡の発達により、細胞に明瞭な核(細胞核)を持たない生物(原核生物)と、細胞に明瞭な核を持つ生物(真核生物)の重要な違いが明らかになった。1937年、エドゥアール・シャットンはこれらの生物を区別するために「原核生物(prokaryote)」と「真核生物(eukaryote)」という用語を導入した。
1938年、ハーバート・F・コープランドは、原核生物からなる新しいモネラ界を創設し、四界の分類を提案した。これは原生生物界のモネラ門を改訂したもので、現在では細菌と古細菌に分類されている生物が含まれていた。エルンスト・ヘッケルは、1904年の著書『The Wonders of Life(生命の不思議)』の中で、藍藻類(またはPhycochromacea)をモネラ門に分類した。これは次第に受け入れられ、藍藻はシアノバクテリア門の細菌として分類されるようになった。
1960年代、ロジャー・スタニエとC.B.ヴァン・ニールは、エドゥアール・シャットンの初期の研究、特に1962年の論文「The Concept of a Bacterium(バクテリアの概念)」を取り上げ、普及させた。これによって初めて、原核生物と真核生物の2帝系より上位の階級、すなわち上界(superkingdom)あるいは帝国(empire)が創設された。後に2帝系は、古細菌、細菌、真核生物の3ドメイン系に拡張される。
五界
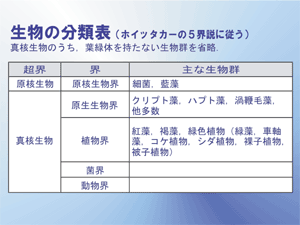
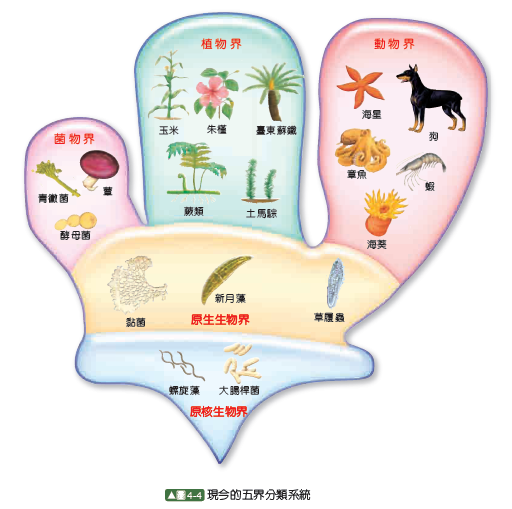
真菌類と、植物とみなされる他の生物との違いは、以前から一部の人々により認識されていた。ヘッケルは、当初の分類の後、真菌類を植物界から原生生物界に移動したが、この分割は、当時の科学者からほとんど無視されていた。ロバート・ホイッタカーは、真菌類のための界(Fungi)の創設を認めた。その結果、1969年にホイッタカーによって提案された五界分類系は一般的な標準となり、今でも改良を加えながら多くの著作で使用され、新しい多界分類系の基礎となっている。これは主に栄養の違いに基づいており、植物界のほとんどは多細胞の独立栄養生物で、動物界は多細胞の従属栄養生物で、そして菌界は多細胞の腐生生物であった。
残りの2つの界、原生動物とモネラには、単細胞コロニーと単純型細胞コロニーが含まれていた。五界系は2帝系と組み合わされることもある。ホイッタカーの体系では植物界は一部の藻類を含んでいた。リン・マーギュリスの五界系のような別の体系では、植物には陸上植物(有胚植物、Embryophyta)だけを含み、原生生物界はより広い定義を持っている。
ホイッタカーの体系が発表された後、五界モデルは高校の生物学の教科書で広く使われるようになった。しかし、ほとんどの科学者が二界系から五界系へと移行したにもかかわらず、1975年になっても一部の著者は伝統的な動物と植物の2界体系を使い続け、植物界を原核生物門(細菌とシアノバクテリア)、真菌類門(真菌類とその近縁種)、緑藻類門(藻類と陸上植物)の亜界(subkingdoms)に分割した。
六界
1977年、カール・ウーズらは、リボソームRNAの遺伝子構造に基づいて、原核生物を真正細菌(Eubacteria、後に細菌/Bacteriaと呼ばれる)と古細菌(Archaebacteria、後のArchaea)に基本的に分類することを提案した。これは後に、細菌、古細菌、真核生物という「生命の3ドメイン」の提案につながる。これと五界モデルを組み合わせることで、モネラ界を菌界(kingdom Bacteria)と古細菌界(kingdom Archaea)に置き換えた六界モデルが作られた。この六界モデルは、最近では米国の高校でも生物学の教科書に使われているが、現在の科学的コンセンサスを危うくするという批判もある。しかし、原核生物を2つの界に分けることは、主に原生動物(Protista)が原生動物(Protozoa)とクロミスタ(Chromista)に置き換えた点で異なるものの、トーマス・キャバリエ=スミスによる最近の七界説でもまだ使われている。
八界
トーマス・キャバリエ=スミスは、真正細菌(Eubacteria)と古細菌(Archaebacteria)の違いは非常に大きく(特にリボソームRNA遺伝子の遺伝的距離を考慮すると)、原核生物を2つの異なる界に分離する必要があるという当時のコンセンサスを支持した。そして真正細菌を2つの亜界、ネギバクテリア(グラム陰性細菌)とポジバクテリア(グラム陽性細菌)に分けた。電子顕微鏡の技術的進歩により、植物界からクロミスタを分離することが可能になった。実際、クロミスタの葉緑体は(細胞質ではなく)小胞体の内腔に位置している。さらに、クロミスタだけがクロロフィルcを含む。それ以来、二次的に葉緑体を失ったと考えられる多くの非光合成的な原生生物門が、クロミスタ界に統合された。
やがてついに、ミトコンドリアを持たない原生生物が発見された。ミトコンドリアはプロテオバクテリアの内部共生の結果であることが知られていたため、これらのミトコンドリアを欠く真核生物は、真核形成の重要な一歩を踏み出した原始的なものであると考えられた。その結果、これらのミトコンドリアを欠く原生生物は原生生物界から分離され、上界および界としてアーケゾア(kingdom Archezoa)が同時に誕生した。この上界は、他の5つの真核生物界(動物界、原生動物界、菌界、植物界、クロミスタ界)をまとめたメタカリオタ上界(Metakaryota)と対立していた。このアーケゾア仮説は後に放棄され、後の仮説にはアーケゾアとメタカリオタの区分を含めなかった。
‡分類学者にはもう認識されていない。
六界(1998)
1998年、キャバリエ=スミスは六界モデルを発表したが、その後の論文で改訂されている。次に、2009年に発表された改訂版を説明する。キャバリエ=スミスは、ウーズらによって提唱され、最近の研究によって裏付けられている基本的な真正細菌(Eubacteria)と古細菌(Archaebacteria)を区別することの重要性を否定している。細菌界(原核生物帝の唯一の界)は膜トポロジーによって、ウニバクテリア(Unibacteria)とネギバクテリア(Negibacteria、グラム陰性細菌)の2つの亜界に分割された。ウニバクテリアは古細菌(Archaebacteria)とポジバクテリア(Posibacteria、グラム陽性細菌)に分けられた。古細菌の遺伝的距離の長い枝よりも、二膜性-単膜性を区別する方がはるかに本質的であり、これには特に生物学的意義はないと見なすものであった。
キャバリエ=スミスは、分類群は単系統でなければならないという要件を有効とは認めていない。彼は、原核生物、細菌、ネギバクテリア、ウニバクテリア、ポジバクテリアを有効な側系統分類群として定義し、(生物学的ニッチの概念に関して)生物学的意義の重要な革新を示している。
同じように、彼の側系統的な原生動物界には、動物界、菌界、植物界、クロミスタのそれぞれの祖先が含まれている。系統発生学的な研究の進歩により、キャバリエ=スミスは、アーケゾア(すなわちミトコンドリアを持たない原始的な真核生物)であると考えられていたすべての系統が、実際には、ミトコンドリアを二次的に失い、典型的にはミトコンドリアを新しい細胞小器官であるハイドロジェノソームに変化させていたことに気づいた。つまり、キャバリエ=スミスによって与えられた用語の意義によれば、すべての生きている真核生物は実際にはメタカリオタである。微胞子虫門(phylum Microsporidia)のように、消滅したアーケゾア界の一部は、菌界に再分類された。他には、現在エクスカバータ下界に属しているメタモナーダのように、原生動物界に再分類されたものもある。
キャバリエ=スミスは側系統を認めているので、下の図は「祖先図」ではなく「組織図」であり、進化系統樹を表しているわけではない。
七界
キャバリエ=スミスらは、2015年に分類を改訂した。彼らはこの枠組みで、原核生物と真核生物の2つの上界と、その下に7つの界を導入した。原核生物には細菌界と古細菌界という2つの界がある。(これは「細菌と古細菌の分類学的概要」と「生命のカタログ」に示されたコンセンサスに基づいている)。真核生物には、原生動物界、クロミスタ界、植物界、菌界、動物界の5つの界がある。この分類のもとで、原生生物(protist)は、真核の単細胞生物のどれかに位置づけられる。
まとめ
生物の界レベルの分類は、(この手法にはいくつかの問題があるにもかかわらず)生物をグループ分けする有用な方法として今でも広く使われている。
- 原生動物界(Protozoa)のような界は、クレードというよりむしろ段階を表しているため、系統分類システムでは否定されている。
- 最新の研究は、真核生物をどの標準的な分類体系に分類することも支持していない。2010年の時点では、どの界も研究によって十分に裏付けられておらず、広く受け入れられるには至っていない。2009年、アンドリュー・ロジャーとアラステア・シンプソンは、新発見を分析する際には慎重であるべきと強調し、「真核生物の生命樹に関する我々の理解における現在の変化のペースを考えると、我々は慎重に進まなければならない」と語った。
伝統的な界の後継
界という概念は一部の分類学者によって使用され続けているものの、生物を自然のグループに並べることに重点を置く分岐分類を提供するものとは見なされておらず、伝統的な界から脱却する動きがある。
生命の3ドメイン
1970年代半ば以降、分類の主な要因として分子レベルの遺伝子(当初はリボソームRNA遺伝子)の比較が重視されるようになり、外見や行動よりも遺伝子の相同性が強調されるようになった。界を含む分類階級は、単系統群(共通祖先の「すべての子孫」)であれ、側系統群(共通祖先の「一部の子孫」のみ)であれ、共通の祖先を持つ生物のグループとされた。
このようなRNAの研究に基づいて、カール・ウーズは、生命は3つの大きな区分に分けられると考え、それらを「3超界(three primary kingdom)」モデルあるいは「超界(urkingdom)」モデルと呼んだ。
ウーズらは、1990年、最高位の名称として「ドメイン」を提案した。この用語は、1974年にムーアによって導入された「ドミニオン」(dominion、羅: dominium)というカテゴリーの同義語である。ムーアと異なるのは、1990年にウーズらがこのカテゴリに対するラテン語を提案していなかったことで、的確に導入された用語であるドミニオンが支持される論拠となっている。
ウーズは原核生物(以前はモネラ界に分類されていた)を真正細菌と古細菌という2つのグループに分け、この2つのグループの間にある遺伝的な差異は、そのどちらか一方と、すべての真核生物の間にある差異と同じくらい多くあることを強調した。
遺伝子データによれば、植物、真菌類、動物などの真核生物グループは、見た目は異なるかもしれないが、真正細菌や古細菌よりも互いに近縁である。また、真核生物は真正細菌よりも古細菌に近いこともわかった。真正細菌と古細菌とを区別する優位性は疑問視されてきたが、その後の研究によって支持されてきた。ウーズが提唱した分類体系にいくつの界が存在するかについては、コンセンサスを得ていない。
真核生物のスーパーグループ
2004年、シンプソンとロジャーは総説論文で、原生生物は「動物、植物、真菌類以外のすべての種々雑多な真核生物である」と指摘した。彼らは、分類の正式な階級として認められるのは単系統グループのみに限るべきで、以前はこのアプローチは非現実的だったが(文字どおり数10もの真核生物の界を必要とした)、今では真核生物を「おそらくはすべて単系統である、わずか数個の主要なグループ」に分けることが可能になったと主張した。
これに基づいて作られた真核生物の実際の「界」(引用符は彼らによる)を右側の図に示す。2005年、この方法を用いた分類が、国際原生生物学会に向けて「多くの学会の専門家と協力した」委員会によって作成された。ここでは、真核生物を同じ6つの「スーパーグループ」に分割した。発表された分類では「界」を含む正式な分類階級を意図的に使用していない。
このシステムでは、多細胞動物(Metazoa、後生動物)は、単細胞の襟鞭毛虫や真菌類と同じく、オピストコンタを形成する祖先の子孫である。植物は、動物と真菌よりも遠い関係にあると考えられている。
しかし、国際原生生物学会の分類が発表された同じ年(2005年)には、これらのスーパーグループの一部、特にクロムアルベオラータ(Chromalveolata)が単系統であるかどうかについて疑問が表明され、2006年の再調査では、提案された6つのスーパーグループのいくつかに証拠がないことが指摘された。
2010年現在、リザリア(Rhizaria)はストラメノパイル(Stramenopiles)およびアルベオラータ(Alveolata)と一緒にSARスーパーグループと呼ばれるクレードに属し、リザリアは主要な真核生物グループの1つではないという見解が広まっている。それ以上のコンセンサスはないようである。Rogozinらは、2009年、「真核生物の深い系統分類は、極めて困難で議論の多い問題である」と指摘した。2010年の時点で、2005年に提唱された6つのスーパーグループモデルは、真核生物の真の系統を反映しておらず、したがって真核生物はどのように分類されるべきかという点では合意が得られているようだが、どのようなモデルがそれに代わるべきかという点での合意は得られていない。
最上位の分類の比較
著者の中には、分類に非細胞生物を加えたものもいる。これによって、非細胞生命の Acytota(Aphanobionta とも呼ばれる)というスーパードメインが作られ、もう1つのスーパードメインが細胞生命である Cytota によって構成される。エオサイト説では、真核生物は古細菌の中のテルモプロテオータ(Thermoproteota、以前はエオサイト(eocytes)またはクレン古細菌門(Crenarchaeota)として知られていた)と呼ばれる門から出現したと提案している。
ウイルス
国際ウイルス分類委員会は、ウイルスの分類に「界(kingdom)」という分類階級(接尾辞 -virae が付く)を使用しているが、これはレルムとサブレルムという最上位分類よりも下に位置する。
ウイルスが生命樹に含まれるかどうかについては、現在も議論が続いている。反対の論拠としては、ウイルスは代謝を欠き、宿主細胞の外で複製することができない偏性細胞内寄生生物であるという事実がある。あるいは、ウイルスにはさまざまな進化的起源があると考えられており、宿主からヌクレオチド配列を取り入れる傾向をもつため、系統樹に含めるのは問題があるという意見もある。
その一方では、ウイルスを含めることに賛成する論拠もある。そのひとつが、典型的な細胞遺伝子を持つミミウイルス(Mimivirus)のような、異常に大きく複雑なウイルスの発見からきている。
注釈
出典
推薦文献
- Raven, Peter H.; Evert, Ray Franklin; Eichhorn, Susan E. (2013). Biology of plants (Eighth edition ed.). New York: W.H. Freeman and Company Publishers. ISBN 978-1-4292-1961-7. OCLC 781446671. https://www.worldcat.org/title/781446671 (英語)
- Pelentier, B. (2007-2015). “Empire Biota: a comprehensive taxonomy”. 2014年2月2日時点のオリジナルよりアーカイブ。2015年10月11日閲覧。- 歴史的概要(英語)
関連項目
- 分岐分類学 - 生物をクレードに分類する生物学的な分類法
- 系統学 - 生物のグループ間またはグループ内の歴史と関係を研究する学問
- 体系学 - 生物の多様化および生物間の時間的な関係を研究する学問
- 分類学 - 共通の特徴に基づいて生物群を命名/定義/分類する科学的研究
- 生物の分類 - 共通の特徴に基づいて生物群を命名し、定義し、分類する科学的研究
- 階級 (生物学) - 祖先または遺伝的階層における生物群(分類群)の相対的な水準
- リンネ式階層分類体系 - カール・リンネによる生物学的分類体系(1735年)、または伝統的な意味での分類学